

Juliane Rocio Neves1; Fábio Francesconi2; Adilson Costa3; Beatriz de Medeiros Ribeiro4; Ivonise Follador5; Luiz Maurício Costa Almeida6
Propionibacterium acnes (P. acnes) is one of the main microorganisms found on the skin. It is predominantly found in hair follicles, prefers anaerobic conditions, preferably colonizes the areas with high sebum production, and is the main bacterium involved in the pathogenesis of acne. The indiscriminate use of antibiotics for the treatment of acne vulgaris can result in the development of bacterial resistance. The present article is aimed at updating dermatologists with the most current data - from the classification to the physiopathogenic mechanisms involved in bacterial resistance to P. acnes and its possible clinical implications.
Keywords: PROPIONIBACTERIUM ACNES; DRUG RESISTANCE, BACTERIAL; ACNE VULGARIS
Propionibacterium acnes (P. acnes) is a gram-positive, facultative anaerobic, diphtheroid-type, non-spore forming bacillus.1 This bacterium is part of the skin's microbiome and has a confirmed presence in the stratum corneum and pilosebaceous units.2
P. acnes contributes to half of the skin's microbiome, with an estimated density of 102 to 106 clones per square centimeter.3 In the skin, its distribution is prevalent in the facial and scalp areas, and is related to the high concentration of pilosebaceous units in these sites.4 It is common in areas rich in eccrine sweat glands and mucous membranes, however it is also present in small amounts in the lower limbs.5 This agent is also part of the microbiome of the conjunctiva, outer ear, oral cavity, and upper respiratory tract.6 It can be occasionally commensal in the peripheral lung tissue and mediastinal lymph nodes.7
P. acnes was previously called Corynebacterium acnes, in reference to its ability to ferment carbohydrates into propionic acid and light chain fatty acids (LCFA) - substances with known antimicrobial activity.8 Lipids and fatty acids, as well as pantothenate, nicotinamide acids are the main sources of nutrition for this bacterium, in addition to some other elements such as cobalt and iron that make up the diet.9, 10
P. acnes is known for contributing to health by inhibiting the invasion of the skin by common pathogens such as Staphylococcus aureus and Streptococcus pyogenes.11 The hydrolyzation of triglycerides with the release of free fatty acids contributes to the acid pH of the skin's surface, which is another well-known factor for skin protection.12 Fermentation of glycerol has proven to have in vitro and in vivo probiotic action, suppressing the growth of the USA 300 methicillin-resistant Staphylococcus aureus, one of the most prevalent strains in the community.11
The P. acnes genome encodes all the key components for the oxidative phosphorylation and has the genes for the cytochrome c oxidase, which ensures its ability to grow in different metabolic conditions.13-15 Thus, P. acnes is able to "tolerate" exposure to oxygen for a few hours and survive in vitro for up to eight months under anaerobic conditions.16 Many of its genes have recognized virulence factors, conferring pathogenic potential to this bacterium.17
The sub-classification of P. acnes was introduced by Johnson and Cummins with the demonstration of two distinct phenotypes of this agent (Type I and Type II), based on studies of serum agglutination and analysis of the cell wall sugars. 18 Subsequently, the phylogenetic analysis of the RecA gene and hemolysin/cytotoxin (tly) of P. acnes strains proved different strains of both types.19 Type I was further divided into IA and IB,19, 20 and Subtype III was described.21 Genetic division of P. acnes based on multilocus sequence typing technology (MLST), identifies the following subtypes: Type I - IA1, IA2, IB, IC or I-1a, I-1b, I-2, Type II and Type III.22-24
With the sequencing of the P. acnes genome (KPA171202, IB strain) the knowledge of this bacterium has further advanced.25 The analysis evidenced a genome of 2.56Mb with 60% GC, encoding 2,333 open reading frames (ORFs), with multiple products such as sialidases, neuraminidases, endoglycoceramidases/endoglycosidases, lipases, and pore-forming enzymes.25 When 82 strains of P. acnes had their genomes compared, it was possible to identify concordance in 88% of the genome (2.2Mb). Unique single nucleotidepolymorphisms (SNPs) were identified in the central region 122,223, with the possibility of using the ribotyping of the gene 16S in the construction of the phylogenetic tree of P. acnes having been demonstrated.26 The non-central genome, which is not shared by all strains, corresponds to 0.90Mb and assists in distinguishing between different strains.26
Combining the knowledge of the bacterium's genome and the greater ease of use of the technique based on the sequencing of the 16 S rRNA as compared with the MLST technique (the first uses one gene as compared with the second, which uses 6-9), it became possible to define the P. acnes' ribotypes (RT), a fingerprint of the restriction fragments of genomic DNA. The analysis of sixty-nine strains allowed for the identification of the following RTs: 19 RTs1, 5 RTs2, 15 RTs3, 8 RTs4, 7 RTs5, 4 RTs6, 6 RTs8, four strains of smaller RTs, and one III strain.26
On average, each individual has 3±2 P. acnes ribotypes, with three or more clones with no difference in the abundance of the bacteria.3 Among the more frequently found ten ribotypes, RT1, RT2, and RT3 were the most prevalent in patients with and without acne. RT4, RT5 (corresponding to the IA-2 class, which is considered an acne-specific subtype) and RT8 were more prevalent in patients bearing acne. RT6 was found only in healthy patients.3, 26, 27
Knowledge of the genetic diversity of P. acnes made it possible to understand why a ubiquitous bacterium in humans may be directly related to acne in some individuals and still be critical to the microbiome balance.24, 28
Genomic and proteomic analyses complement the current knowledge when it demonstrates different profiles, with four described proteomic patterns. These patterns also differ according to the conditions of the culture medium, with unique patterns in aerobic and anaerobic conditions, suggesting there is influence of the medium on the bacteria.29
P. acnes was historically considered an agent of low pathogen potential, however the current knowledge about these bacteria indicates a genetic diversity with different levels of potential for virulence.26
In addition to the single loci 1, 2, and 3 found in RT4 and RT5 (class IA-2) - subtypes associated with acne - a linear plasmid might have a role in the physio-pathogenesis of acne.26 It is predominantly found in acne lesions.30 Genomic loci seem to be originated from mobile genetic elements that encode virulence genes.31 The RT8 strains of IB-1 class, also related to acne, have a single genomic island (20kb locus encoding non-ribosomal peptide synthetases (NRPS), which are possible virulence factors.4
The RT2 strain belongs to Class II and is distributed both in acne-bearing and healthy patients; however RT6 strain prevails in the latter group.3 The most characteristic genetic feature of these strains is the presence of the clustered regularly interspaced short palindromic repeats locus (CRISPR)/Cas)3. The CRISPR/Cas system provides the bacteria with immunity against viruses and plasmids.32, 33 Bacteriophage infections have been associated with the potential pathogenic of bacteria and is also a possible therapeutic agent.34, 35
Type II strains have decreased lipase activity, which is related to the virulence of P. acnes.10
Type III strains are rarely found on the skin's surface.36 There is a loss of 43kb in the length of the genome with 42ORFs of specific genomic property.26 This subtype is related to vertebral disc infection.27, 37
a) Influence of the metabolism
P. acnes can be related to the initial stage of acne due to the fact that it causes an increase in the lipogenesis originated in sebaceous glands.38 It acts through the action of soluble factors or direct action with an increase in the production of 15-deoxy-12, 14 -prostaglandin J2 (15d-PGJ2) via the cytochrome P450 with increased triacylglycerol synthesis.38, 39 IGF-1 and IGF-1R are a well known target of P. acnes.40
There is also evidence that P. acnes influences the differentiation of keratinocytes via an increase in transglutaminase and cytokeratin,17 and a decrease in the expression levels of cytokeratins 1 and 10.41 Some strains are even capable of increasing the involucrin, the expression of mRNA of cytokeratin 6, and decreasing the levels of expression of cytokeratin 6 and 16.41 The effect on keratinocyte differentiation suggests it has influence on the formation of microcomedones.42
b) Biofilm
The production of a biofilm by Propionibacterium acnes - a glycocalyx polymer, which acts as a biological glue - was evidenced by genomic studies.43 The production of the biofilm is closely related to invasive infections and is considered an important factor in these infections, as demonstrated in 93 isolates.44
The production of biofilm can take place in the follicles45 and has been reported in patients with acne in whom the substance led to inflammation that was not associated with cellular immune response.46 When present in the sebum, this polymer leads to the adhesion of keratinocytes, contributing to the formation of comedones. It is believed that this substance has an influence on the immunogenicity, clinical course, and impact on the therapeutic response of acne pictures.43
c) Proteases
There is evidence suggesting P. acnes has a role in increased expressions of interleukins 1's and 8's mRNA, of tumor necrosis factor alpha, of human beta defensin beta 2, and of metalloproteinases 1, 2, 3, 9 and 13 in keratinocytes by activation of proteases, and of proteins activated receptor (protein-activated receptor 2 PAR-2).47 Metalloproteinases can be related both to the pathophysiology of acne48 and scar formation.49
The Christie, Atkins, Munch-Peterson factor (Camp factor) is a secretory protein with co-hemolytic activity and virulence potential against keratinocytes and macrophages that can be encoded by P. acnes.50 Despite being secreted by P. acnes, the specific role of the Camp factor in the pathogenesis of disorders is still unclear.51
Endogenous porphyrins can be produced by P. acnes with a possible influence on perifollicular inflammation by cytotoxic effect and the stimulus of interleukin 8 production.52 After the follicle's rupture and probably by the action of singlet oxygen, the porphyrins can continue their action due to the fact they promote the development of cytotoxic substances, such as squalene peroxide.53
d) Inflammation
In patients with acne, P. acnes is capable of activating innate immunity via toll-like receptor type 2 (TLR2).54 This activation occurs via cell wall components of the bacterium,55, 56 except for when it is not damaged or inactivated.57 In response to the activation of TLR2, there is production of IL-1a by follicular keratinocytes,58 (a proven role in the comedogenesis), production of nuclear factor kappa-light (NF-kB, primary transcriptional factor of fast action) by activated B cells,59 and production of IL-12 and IL-8 by monocytes.54 When in vitro, P. acnes is capable of inducing the mRNA of the MMP-9 and MMP-1 and the expression of MMP-9 (but not MMP-1), through a mechanism dependent on TLR2.48
The in vitro activation of macrophages is accompanied by increased expression of the synthase gene induced by nitrous oxide (inducible nitric oxide synthase - iNOS) and of the cyclooxygenase-2 gene (COX-2), with an increase in nitrous oxide (NO) and of prostaglandin E2 (PG E2) through a dose-dependent mechanism.60 This activation was also observed in keratinocytes. In the same study there is evidence of the toll-like pathway, for when TLR-2 are blocked with inhibitory antibodies, the increase in the expression of genes is halted.60
Innate immunity can also be activated by pattern-recognition cytoplasmic receptors, and the oligomerization domains, by binding with nucleotides (nucleotide binding oligomerization domain - NOD).61 These receptors are termed NOD-like receptors (NLR) and assist in the identification of microorganisms and molecules with potential for damaging the cell, such as reactive oxygen species (ROS).15 When activated by direct binding , the NLR forms a multiprotein complex with adapter proteins and pro-caspase-1, and is then termed inflammasome.62 Four inflammasomes are described as being capable of identifying bacteria: NLRP1, NLRP3, NLRC4 (also known as IPAF) - all of which belong to the NLR family - and a fourth called AIM2 - that does not belong to this family.63 Their activation takes place after the cleavage of pro-caspase-1 active caspase-1, the protease that processes the pro-interleukin 1b and 18 into mature and active interleukins.22
P. acnes activates the NLRP3 on human monocytes.63 It has been shown that the release of IL-1b by monocytes depends on the phagocytosis of P. acnes.61 Strains with proven ability to invade epithelial cells are of type I in 71% of cases.64, 65 IL-1b is potentially responsible for the induction of in vivo neutrophilic inflammatory response induced by P. acnes.66 The bacterium can induce the formation of caspase-1 in neutrophils, with the generation of additional IL-1b and IL-18.67 The activation of the inflammasome can also activate NF-Kb.65 P. acnes also induces the production of IL-1 in sebocytes.68
P. acnes stimulates the production of genes linked to the Th17 immune response, in addition to stimulating the secretion of IL-17 by CD4+ lymphocytes. Vitamin A, and vitamin D are capable of inhibiting the Th17 differentiation induced by P. acnes.69
With the expanding knowledge about the different P. acnes strains and their influence on innate immunity, the identification of the phylogenetic groups tend to occur not only based on the standard of secreted proteins, but also based on the ability to induce different patterns of immune response.70 Another important aspect of the inflammatory cascade induced by P. acnes is that some are stimulated by dead P. acnes or its components, a fact that should be taken into consideration in the acne bearing patient's therapeutic strategy, using substances with bactericidal and anti-inflammatory actions.71
Knowledge of the impact of this bacterium on the immune system has allowed the use of this agent in various immunomodulatory strategies, especially in veterinary medicine, as described in the following examples: the inactivated bacteria stimulates Th1 and Th2 response in mice;72 it is capable of inducing antitumor response,73 increasing the resistance against viral and parasitic infections, preventing focal segmental glomerulosclerosis; 74 it has antibacterial activity;75-78 in murine model of sepsis it reduces the mortality by 50%;79 it is used in a vaccine that improves the atopic dermatitis murine model;80 in addition to its use aimed at preventing infections81 and preparing vaccines.82
Evasion mechanisms
After having been phagocytized, P. acnes can survive and persist within macrophages, interfering or blocking the maturation pathway of phagosomes.83 P. acnes has an intracellular life cycle, offering a possible niche for the survival and spread of the bacteria.83
P. acnes and human diseases
P. acnes has a proven role in acne, however its importance in other diseases is underestimated.
With the ability of growing in medium with diverse oxygenation conditions (in particular in anaerobic settings), of surviving intracellularly in macrophages with the potential to generate inflammation and producing biofilm, P. acnes can be transferred from its habitat (e.g. the skin's microbiome) to deeper tissues, and with the ability to survive and present pathogenic potential.44 In addition to direct contamination through the skin, infection by P. acnes may result from transient bacteremia.84
Infections by this bacterium usually have an indolent course and are difficult to diagnose for it is a common bacterium and differentiating contaminations from infections is always a challenge - nevertheless the formation of biofilm is used to differentiate these two clinical situations.44 In a study with 522 patients, clinically significant bacteremia of P. acnes occurred in 3.5% of cases, of which 55.6% were classified as nosocomial, 33.3% had a history of previous invasive procedure and 5.9% mortality.85
Sarcoidosis
Sarcoidosis is a systemic granulomatous disorder that affects individuals with genetic susceptibility after exposure to a particular environmental stimulus, with Propionibacterium acnes being a proven cause.86 The correlation between the bacterium and sarcoidosis granuloma was demonstrated by techniques of in situ hybridization,87 immunohistochemistry with monoclonal antibodies against P. acnes,88-90 granuloma based isolation in culture, 91 and the complete sequencing of the sarcoidosis bearing patient's bacterium's genome.92 A murine experimental model was capable of inducing sarcoidosis granuloma formation after the administration of a recombinant protein from active93 or dead94 P. acnes.94 Considering the studies on the influence of P. acnes on sarcoidosis, authors have postulated the hypothesis that a hypersensitivity response, with a change in Th1/Th17 balance can generate the sarcoidosis picture.95 In an interesting case report that identifies P. acnes in sarcoidosis granuloma, the patient was treated with clarithromycin with the complete resolution of the condition88.
Prostate cancer
Recent studies relate the chronic inflammatory state with prostate carcinogenesis due to DNA damage that leads to tissue replication, migration, and angiogenesis.96 There is evidence of P. acnes in prostate tissue with chronic inflammation,97 verified through techniques of monoclonal antibody,98 in situ hybridization with fluorescence,99 and immunohistochemistry.100 The infection's pathway is not yet established, but isolation of P. acnes in urine samples suggests the urethra is a possible channel.101, 102 A murine model of chronic prostatic inflammation was possible after transurethral catheterization of P. acnes.103 Categorizing by type, using the MLST technique, demonstrates that prostate strains do not have a cutaneous origin, contradicting researchers who advocate the possibility of contamination during the collection of prostatectomy material.104 Vimentin appears to be a key determinant factor for prostatic tissue invasion.105 Another study correlates P. acnes' titles with prostate cancer.106
Infection of orthopedic prostheses
P. acnes is one of the agents that causes shoulder prosthesis infection most frequently (second only to S. aureus).107 Despite the small number of the sample, the analysis of 22 shoulder prosthetic infection isolates led to the conclusion that the hemolytic phenotype was the one most associated in this case.108 This infection's prevalence varies from 3.9% to 15.4% and delayed diagnosis can have an impact on the success of the procedure, with loss of the prosthesis, chronic pain, exudate and sepsis having been described.109 Spinal prosthetic infections are also related to P. acnes, with an incidence of 0.2%, however possibly reaching 12.0% depending on the used instrumentation.110-112
Endocarditis
Endocarditis caused by P. acnes is rare and is related to heart valve prostheses.113, 114 There are reports of cases that onset after invasive procedures.85 Due to the subacute oligosymptomatic development, diagnosis is often delayed, entailing valve and perivalvar destruction or abscess formation, with mortality rates reaching roughly 18.7%.115 Little is known about the ideal treatment, and surgery is commonly indicated with an aim at draining the abscess or replacing the valve.115
Central nervous system infection
P. acnes is related to post-operative infections of the central nervous system, and among common procedures are the use of shunts, bone grafts, craniotomy, abscess drainage, and rare descriptions of meningitis cases with no previous history of procedure (8 reported cases).116-119 In half of the cases, symptoms may appear acutely within seven days or subacutely within 14 weeks in the other half, on average. Symptoms are those deemed classic for meningitis, with a cerebrospinal fluid (CSF) profile corresponding to that of the aseptic meningitis with mononuclear pleocytosis.19 Based on reports, penicillin G, chloramphenicol or vancomycin are used with favorable results.120 Optic nerve neuritis121 and abscess formation are described complications.120
Other infections
Breast implant infection,17, 122 acute post-traumatic endophthalmitis,123 discitis and spondylodiscitis,124-130 pericarditis,131 aortic stent endarteritis,132, 133 prosthetic eye infection,134 osteomyelitis,135, 136 surgical wound infection,137 endodontic infection138 and keratitis.139
Related dermatological conditions
Progressive macular hypomelanosis of the trunk,140-142 alopecia143 and Sapho syndrome.144
P. acnes diagnosis
Despite the bacterium having aerotolerant characteristics, the diagnosis of P. acnes should be performed in a culture medium with anaerobic conditions - thioglycolate is the routinely used culture medium.145 When enriched, this culture medium has a low redox potential and assists in the growth of the microorganism145.
There should be a systematic subculture on agar plates even in the absence of turbidity.146 The optimum temperature for growth is 37ºC, and the duration of inoculation should be continued for at least 14 days.147 On the other hand, the culture duration should not be too long, for the probability of contamination by growth of bacteria originating from the normal flora would increase - as a result the maximum waiting period has not been established yet.147 The interpretation of cultures should be performed with caution, especially if there is only a single medium. For greater reliability of the result, growth should be observed in more than one culture medium, always in light of the correlated patient's clinical data and, where possible, of other diagnostic methods, such as histology and molecular diagnostics. 146 Fluorescence with in situ hybridization (Fish) is a technique with potential use in blood cultures. It showed 95% sensitivity and 100% specificity in the diagnosis of infection by P. acnes.148
For the isolation and identification of P. acnes, attention should be paid to the collection methodology, for different techniques will demonstrate anatomically distinct populations: swab and scraping are used to identify the more superficial bacteria; the use of tape with cyanoacrylate targets superficial and infundibular populations; the punch evidences deeper follicular populations and can be used in biofilm research; finally there are direct tissular visualization techniques, such as the Fish technique, immunofluorescence microscopy and immunohistochemical techniques.149
Antibiotic resistance by Propionibacterium acnes
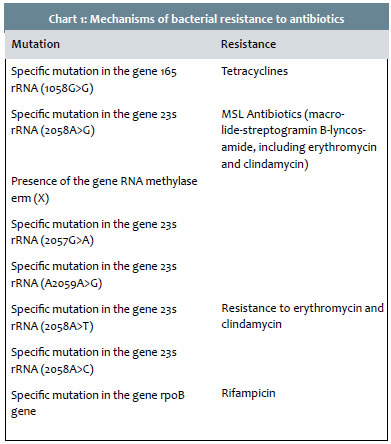
The first documented antibiotic resistance by P. acnes was linked to erythromycin.150 The known mechanisms of resistance are: specific mutations to the genes that encode the ribosomal RNA, single mutation in the 16S rRNA (1058G>C) gene, mutations in the gene 23S rRNA with four recognized phenotypic groups (Group I: 2058A> G, Group II: presence of RNA methylase erm(X) gene in a mobile genetic element, Group III: 2057G> A; Group IV:A2059A> G) and mediated erm(X) resistance.151-152 Resistance to rifampicin is associated with a punctual mutation in the rpoB gene's Clusters I and II, and can be prevented if associated with levofloxacin, clindamycin, and penicillin G153 (Chart 1).
Antibiotic resistance by P. acnes is better studied in acne patients. The most significant resistance profiles for the treatment of acne are associated with specific strains of bacteria, in particular ST3 clone, which has worldwide distribution.3, 22 The ribotypes that are more frequently related to resistance are RT4 and RT5.3 An individual can host a complex population of P. acnes, with a variable number of clones from one to six, which have different pathogen potential and different resistance patterns, issues that directly impact the difficulty in acne treatment. 154 In addition to having the bacteria, a person can transmit the different clones, and thus resistant bacteria can be spread in the population.155
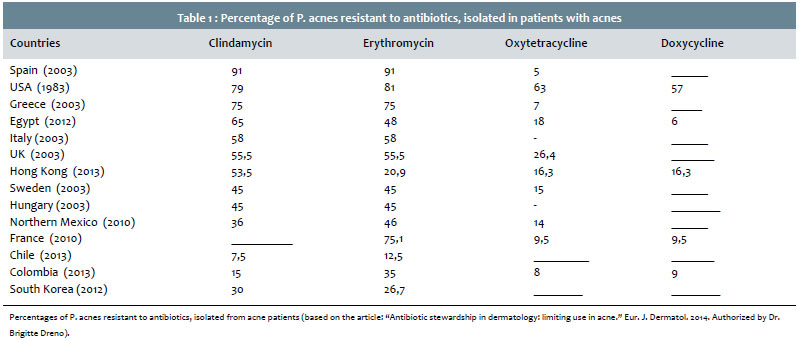
Percentages of P. acnes resistant to antibiotics, isolated from acne patients (based on the article: "Antibiotic stewardship in dermatology: limiting use in acne." Eur. J. Dermatol. 2014. Authorized by Dr. Brigitte Dreno).
There has been an increasing number of cases of antibiotic-resistance by P. acnes over the years: in the UK, the resistance rate increased from 34.5% in 1991 to 55.5% in 2000;156 94% of the isolates in Spain and 51% from the isolates in Hungary were resistant to at least one antibiotic.155 The highest resistance rates are related to erythromycin, with cross-resistance to clindamycin. The lowest rates are linked to tetracyclines.157, 158
The evaluation of 114 isolates in Denmark (72 acne bearing and 42 healthy patients) showed 34% of antibiotic resistance by P. acnes, with 15.8% resistance to clindamycin, 8.8% resistance to erythromycin, and 9.6% resistance to tetracycline, with 39 patients having been affected: 25 acne bearing patients who had the highest proportion of isolates with resistance to tetracycline.154 In Japan, the rate of resistance to macrolides was 4% between 1994 and 1995, however it became much higher and related to the erm(X) gene in 2008.159
Data published in Latin America show a resistance ratio of 33.7% in Chile: 26.3% to trimethoprim-sulfamethoxazole, 12.5% to erythromycin, and 7.5% to clindamycin, with a total cross-resistance between clindamycin and erythromycin, and 40% cross-resistance between erythromycin and trimethoprim-sulfamethoxazole, without identifying resistance to tetracycline and doxycycline.160 The main risk factors for the occurrence of resistance were: older age, previous use of topical antibiotic and, in the case of trimethoprim-sulfamethoxazole, the severity of the acne.160 In Colombia, there were 35% of strains resistant to erythromycin, 15% to clindamycin, 9% to doxycycline, 8% to tetracycline and 1% to minocycline, with 12% cross-resistance between erythromycin and clindamycin, 6% between doxycycline and tetracycline, with previous use of antibiotics being the main risk factor.161 In Mexico there was an 82% resistance to azithromycin, 68% resistance to trimethoprim-sulfamethoxazole, and 46% resistance to erythromycin.162
Elsewhere, in Hong Kong, 54.8% of strains were resistant (53.5% to clindamycin, 20.9% to erythromycin, 16.3% to tetracycline, 16.3% to doxycycline and 16.3% to minocycline), with an 11.6% cross-resistance between clindamycin and erythromycin, and 16.4% of multiple resistors.163 In Egypt, resistance to clindamycin was identified in 66.3%, to erythromycin in 49%, to oxytetracycline in 26.5%, to doxycycline in 16.3%, and to azithromycin in 9.2%.164 (Table 1).
In Australia, growth in the P. acnes resistance rate was not observed between 1997-1998 and 2007.165 Prior use of antibiotics (oral or local) was considered a risk factor for resistance, and the use of retinoids, despite its decreasing effect in the growth rate of P. acnes, did not influence the incidence of resistant strains.165
A European study of 304 isolates of P. acnes from 13 laboratories in 13 different countries tested six antibiotics.166 Blood was the most common source, followed by skin infections, in soft tissue and abdominal infection.166 Of the isolates, 2.6% were resistant to tetracycline, 15.1% to clindamycin, and 17.1% to erythromycin, without descriptions of resistance to linezolid, benzathine penicillin, and vancomycin.166 There was a variation in the resistance profile among countries, with 83% in Croatia, 60% in Italy, and none in Norway, with blood isolates showing a prevalence among the resistant ones.166
As P. acnes has a low susceptibility to cephalosporins, use of antibiotic prophylaxis in surgical procedures (where this bacterium is an important source of post-operative complications) should be revised.167
In cases of severe infections caused by P. acnes, drugs with the possibility of resistance cannot be used, and surgical procedures should be associated with clinical treatment, with a preference - when in combination - for using crystalline penicillin, vancomycin, daptomycin, and rifampicin due to their effect on the biofilm.166, 168, 169
1. Zhang Y, Ertl HC. The effect of adjuvanting cancer vaccines with herpes simplex virus glycoprotein D on melanoma-driven CD8+ T cell exhaustion. J Immunol. 2014;193(4):1836-46.
2. Jahns AC, Oprica C, Vassilaki I, Golovleva I, Palmer RH, Alexeyev OA. Simultaneous visualization of Propionibacterium acnes and Propionibacterium granulosum with immunofluorescence and fluorescence in situ hybridization. Anaerobe. 2013;23:48-54.
3. Fitz-Gibbon S, Tomida S, Chiu BH, Nguyen L, Du C, Liu M, et al. Propionibacterium acnes strain populations in the human skin microbiome associated with acne. J Invest Dermatol. 2013;133(9):2152-60.
4. Beylot C, Auffret N, Poli F, Claudel JP, Leccia MT, Del Giudice P, et al. Propionibacterium acnes: an update on its role in the pathogenesis of acne. J Eur Acad Dermatol Venereol. 2014;28(3):271-8.
5. McGinley KJ, Webster GF, Ruggieri MR, Leyden JJ. Regional variations in density of cutaneous propionibacteria: correlation of Propionibacterium acnes populations with sebaceous secretion. J Clin Microbiol. 1980;12(5):672-5.
6. Brook I, Frazier EH. Infections caused by Propionibacterium species. Rev Infect Dis. 1991;13(5):819-22.
7. Ishige I, Eishi Y, Takemura T, Kobayashi I, Nakata K, Tanaka I, et al. Propionibacterium acnes is the most common bacterium commensal in peripheral lung tissue and mediastinal lymph nodes from subjects without sarcoidosis. Sarcoidosis Vasculitis and Diffuse Lung Diseases. 2005;22(1):33-42.
8. Higaki S, Nakamura M, Morohashi M, Yamagishi T. Propionibacterium acnes biotypes and susceptibility to minocycline and Keigai-rengyo-to. Int J Dermatol. 2004;43(2):103-7.
9. Holland KT, Greenman J, Cunliffe WJ. Growth of cutaneous propionibacteria on synthetic medium; growth yields and exoenzyme production. J Appl Bacteriol. 1979;47(3):383-94.
10. Ferguson DA, Jr., Cummins CS. Nutritional requirements of anaerobic coryneforms. J Bacteriol. 1978;135(3):858-67.
11. Shu M, Wang Y, Yu J, Kuo S, Coda A, Jiang Y, et al. Fermentation of Propionibacterium acnes, a commensal bacterium in the human skin microbiome, as skin probiotics against methicillin-resistant Staphylococcus aureus. PLoS One. 2013;8(2):e55380.
12. Grice EA, Segre JA. The skin microbiome. Nat Rev Microbiol. 2011;9(4):244-53.
13. Bruggemann H. Insights in the pathogenic potential of Propionibacterium acnes from its complete genome. Semin Cutan Med Surg. 2005;24(2):67-72.
14. Gribbon EM, Shoesmith JG, Cunliffe WJ, Holland KT. The microaerophily and photosensitivity of Propionibacterium acnes. J Appl Bacteriol. 1994;77(5):583-90.
15. Cove JH, Holland KT, Cunliffe WJ. Effects of oxygen concentration on biomass production, maximum specific growth rate and extracellular enzyme production by three species of cutaneous propionibacteria grown in continuous culture. J Gen Microbiol. 1983;129(11):3327-34.
16. Csukas Z, Banizs B, Rozgonyi F. Studies on the cytotoxic effects of Propionibacterium acnes strains isolated from cornea. Microb Pathog. 2004;36(3):171-4.
17. Portillo ME, Corvec S, Borens O, Trampuz A. Propionibacterium acnes: an underestimated pathogen in implant-associated infections. BioMed research international. 2013;2013:804391.
18. Johnson JL, Cummins CS. Cell wall composition and deoxyribonucleic acid similarities among the anaerobic coryneforms, classical propionibacteria, and strains of Arachnia propionica. J Bacteriol. 1972;109(3):1047-66.
19. Valanne S, McDowell A, Ramage G, Tunney MM, Einarsson GG, O'Hagan S, et al. CAMP factor homologues in Propionibacterium acnes: a new protein family differentially expressed by types I and II. Microbiology. 2005;151(Pt 5):1369-79.
20. McDowell A, Valanne S, Ramage G, Tunney MM, Glenn JV, McLorinan GC, et al. Propionibacterium acnes types I and II represent phylogenetically distinct groups. J Clin Microbiol. 2005;43(1):326-34.
21. McDowell A, Perry AL, Lambert PA, Patrick S. A new phylogenetic group of Propionibacterium acnes. J Med Microbiol. 2008;57(Pt 2):218-24.
22. McDowell A, Barnard E, Nagy I, Gao A, Tomida S, Li H, et al. An expanded multilocus sequence typing scheme for propionibacterium acnes: investigation of 'pathogenic', 'commensal' and antibiotic resistant strains. PLoS One. 2012;7(7):e41480.
23. McDowell A, Gao A, Barnard E, Fink C, Murray PI, Dowson CG, et al. A novel multilocus sequence typing scheme for the opportunistic pathogen Propionibacterium acnes and characterization of type I cell surface-associated antigens. Microbiology. 2011;157(Pt 7):1990-2003.
24. Lomholt HB, Kilian M. Population genetic analysis of Propionibacterium acnes identifies a subpopulation and epidemic clones associated with acne. PLoS One. 2010;5(8):e12277.
25. Bruggemann H, Henne A, Hoster F, Liesegang H, Wiezer A, Strittmatter A, et al. The complete genome sequence of Propionibacterium acnes, a commensal of human skin. Science. 2004;305(5684):671-3.
26. Tomida S, Nguyen L, Chiu BH, Liu J, Sodergren E, Weinstock GM, et al. Pan-genome and comparative genome analyses of propionibacterium acnes reveal its genomic diversity in the healthy and diseased human skin microbiome. MBio. 2013;4(3):e00003-13.
27. McDowell A, Nagy I, Magyari M, Barnard E, Patrick S. The opportunistic pathogen Propionibacterium acnes: insights into typing, human disease, clonal diversification and CAMP factor evolution. PLoS One. 2013;8(9):e70897.
28. Higaki S, Kitagawa T, Kagoura M, Morohashi M, Yamagishi T. Correlation between Propionibacterium acnes biotypes, lipase activity and rash degree in acne patients. J Dermatol. 2000;27(8):519-22.
29. Dekio I, Culak R, Fang M, Ball G, Gharbia S, Shah HN. Correlation between phylogroups and intracellular proteomes of Propionibacterium acnes and differences in the protein expression profiles between anaerobically and aerobically grown cells. Biomed Res Int. 2013;2013:151797.
30. Kwon HH, Yoon JY, Park SY, Suh DH. Analysis of distribution patterns of Propionibacterium acnes phylotypes and Peptostreptococcus species from acne lesions. Br J Dermatol. 2013;169(5):1152-5.
31. Kasimatis G, Fitz-Gibbon S, Tomida S, Wong M, Li H. Analysis of complete genomes of Propionibacterium acnes reveals a novel plasmid and increased pseudogenes in an acne associated strain. Biomed Res Int. 2013;2013:918320.
32. Horvath P, Barrangou R. CRISPR/Cas, the immune system of bacteria and archaea. Science. 2010;327(5962):167-70.
33. Garneau JE, Dupuis ME, Villion M, Romero DA, Barrangou R, Boyaval P, et al. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 2010;468(7320):67-71.
34. Bruggemann H, Lood R. Bacteriophages infecting Propionibacterium acnes. Biomed Res Int. 2013;2013:705741.
35. Marinelli LJ, Fitz-Gibbon S, Hayes C, Bowman C, Inkeles M, Loncaric A, et al. Propionibacterium acnes bacteriophages display limited genetic diversity and broad killing activity against bacterial skin isolates. mBio. 2012;3(5): e00279-12.
36. Dekio I, Rajendram D, Morita E, Gharbia S, Shah HN. Genetic diversity of Propionibacterium acnes strains isolated from human skin in Japan and comparison with their distribution in Europe. J Med Microbiol. 2012;61(Pt 5):622-30.
37. Mak TN, Sfanos KS, Bruggemann H. Draft Genome Sequences of Two Strains of Propionibacterium acnes Isolated from Radical Prostatectomy Specimens. Genome announcements. 2013;1(6): e01071-13.
38. Iinuma K, Sato T, Akimoto N, Noguchi N, Sasatsu M, Nishijima S, et al. Involvement of Propionibacterium acnes in the augmentation of lipogenesis in hamster sebaceous glands in vivo and in vitro. J Invest Dermatol. 2009;129(9):2113-9.
39. Iwata C, Akimoto N, Sato T, Morokuma Y, Ito A. Augmentation of lipogenesis by 15-deoxy-Delta12,14-prostaglandin J2 in hamster sebaceous glands: identification of cytochrome P-450-mediated 15-deoxy-Delta12,14-prostaglandin J2 production. J Invest Dermatol. 2005;125(5):865-72.
40. Isard O, Knol AC, Aries MF, Nguyen JM, Khammari A, Castex-Rizzi N, et al. Propionibacterium acnes activates the IGF-1/IGF-1R system in the epidermis and induces keratinocyte proliferation. J Invest Dermatol. 2011;131(1):59-66.
41. Akaza N, Akamatsu H, Kishi M, Mizutani H, Ishii I, Nakata S, et al. Effects of Propionibacterium acnes on various mRNA expression levels in normal human epidermal keratinocytes in vitro. J Dermatol. 2009;36(4):213-23.
42. Jarrousse V, Castex-Rizzi N, Khammari A, Charveron M, Dreno B. Modulation of integrins and filaggrin expression by Propionibacterium acnes extracts on keratinocytes. Archives for dermatological research Archiv fur dermatologische Forschung. 2007;299(9):441-7.
43. Burkhart CG, Burkhart CN. Expanding the microcomedone theory and acne therapeutics: Propionibacterium acnes biofilm produces biological glue that holds corneocytes together to form plug. J Am Acad Dermatol. 2007;57(4):722-4.
44. Holmberg A, Lood R, Morgelin M, Soderquist B, Holst E, Collin M, et al. Biofilm formation by Propionibacterium acnes is a characteristic of invasive isolates. Clin Microbiol Infect. 2009;15(8):787-95.
45. Jahns AC, Lundskog B, Ganceviciene R, Palmer RH, Golovleva I, Zouboulis CC, et al. An increased incidence of Propionibacterium acnes biofilms in acne vulgaris: a case-control study. Br J Dermatol. 2012;167(1):50-8.
46. Alexeyev OA, Lundskog B, Ganceviciene R, Palmer RH, McDowell A, Patrick S, et al. Pattern of tissue invasion by Propionibacterium acnes in acne vulgaris. Journal of dermatological science. 2012;67(1):63-6.
47. Lee SE, Kim JM, Jeong SK, Jeon JE, Yoon HJ, Jeong MK, et al. Protease-activated receptor-2 mediates the expression of inflammatory cytokines, antimicrobial peptides, and matrix metalloproteinases in keratinocytes in response to Propionibacterium acnes. Archives for dermatological research Archiv fur dermatologische Forschung. 2010;302(10):745-56.
48. Jalian HR, Liu PT, Kanchanapoomi M, Phan JN, Legaspi AJ, Kim J. All-trans retinoic acid shifts Propionibacterium acnes-induced matrix degradation expression profile toward matrix preservation in human monocytes. J Invest Dermatol. 2008;128(12):2777-82.
49. Sato T, Kurihara H, Akimoto N, Noguchi N, Sasatsu M, Ito A. Augmentation of gene expression and production of promatrix metalloproteinase 2 by Propionibacterium acnes-derived factors in hamster sebocytes and dermal fibroblasts: a possible mechanism for acne scarring. Biol Pharm Bull. 2011;34(2):295-9.
50. Nakatsuji T, Tang DC, Zhang L, Gallo RL, Huang CM. Propionibacterium acnes CAMP factor and host acid sphingomyelinase contribute to bacterial virulence: potential targets for inflammatory acne treatment. PLoS One. 2011;6(4):e14797.
51. Liu PF, Nakatsuji T, Zhu W, Gallo RL, Huang CM. Passive immunoprotection targeting a secreted CAMP factor of Propionibacterium acnes as a novel immunotherapeutic for acne vulgaris. Vaccine. 2011;29(17):3230-8.
52. Schaller M, Loewenstein M, Borelli C, Jacob K, Vogeser M, Burgdorf WH, et al. Induction of a chemoattractive proinflammatory cytokine response after stimulation of keratinocytes with Propionibacterium acnes and coproporphyrin III. Br J Dermatol. 2005;153(1):66-71.
53. Saint-Leger D, Bague A, Lefebvre E, Cohen E, Chivot M. A possible role for squalene in the pathogenesis of acne. II. In vivo study of squalene oxides in skin surface and intra-comedonal lipids of acne patients. Br J Dermatol. 1986;114(5):543-52.
54. Kim J, Ochoa MT, Krutzik SR, Takeuchi O, Uematsu S, Legaspi AJ, et al. Activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. J Immunol. 2002;169(3):1535-41.
55. Ingham E, Walters CE, Eady EA, Cove JH, Kearney JN, Cunliffe WJ. Inflammation in acne vulgaris: failure of skin micro-organisms to modulate keratinocyte interleukin 1 alpha production in vitro. Dermatology. 1998;196(1):86-8.
56. Walters CE, Ingham E, Eady EA, Cove JH, Kearney JN, Cunliffe WJ. In vitro modulation of keratinocyte-derived interleukin-1 alpha (IL-1 alpha) and peripheral blood mononuclear cell-derived IL-1 beta release in response to cutaneous commensal microorganisms. Infect Immun. 1995;63(4):1223-8.
57. Graham GM, Farrar MD, Cruse-Sawyer JE, Holland KT, Ingham E. Proinflammatory cytokine production by human keratinocytes stimulated with Propionibacterium acnes and P. acnes GroEL. Br J Dermatol. 2004;150(3):421-8.
58. Selway JL, Kurczab T, Kealey T, Langlands K. Toll-like receptor 2 activation and comedogenesis: implications for the pathogenesis of acne. BMC dermatology. 2013;13:10.
59. Takeda K, Kaisho T, Akira S. Toll-like receptors. Ann Rev Immunol. 2003;21:335-76.
60. Tsai HH, Lee WR, Wang PH, Cheng KT, Chen YC, Shen SC. Propionibacterium acnes-induced iNOS and COX-2 protein expression via ROS-dependent NF-kappaB and AP-1 activation in macrophages. Journal of dermatological science. 2013;69(2):122-31.
61. Contassot E, French LE. New insights into acne pathogenesis: propionibacterium acnes activates the inflammasome. J Invest Dermatol. 2014;134(2):310-3.
62. Qin M, Pirouz A, Kim MH, Krutzik SR, Garban HJ, Kim J. Propionibacterium acnes Induces IL-1beta Secretion via the NLRP3 Inflammasome in Human Monocytes. J Invest Dermatol. 2014;134(6):1779.
63. Kistowska M, Gehrke S, Jankovic D, Kerl K, Fettelschoss A, Feldmeyer L, et al. IL-1beta drives inflammatory responses to propionibacterium acnes in vitro and in vivo. J Invest Dermatol. 2014;134(3):677-85.
64. Furukawa A, Uchida K, Ishige Y, Ishige I, Kobayashi I, Takemura T, et al. Characterization of Propionibacterium acnes isolates from sarcoid and non-sarcoid tissues with special reference to cell invasiveness, serotype, and trigger factor gene polymorphism. Microb Pathog. 2009;46(2):80-7.
65. Tanabe T, Ishige I, Suzuki Y, Aita Y, Furukawa A, Ishige Y, et al. Sarcoidosis and NOD1 variation with impaired recognition of intracellular Propionibacterium acnes. Biochim Biophys Acta. 2006;1762(9):794-801.
66. Kistowska M, Meier B, Proust T, Feldmeyer L, Cozzio A, Kuendig T, et al. Propionibacterium acnes Promotes Th17 and Th17/Th1 Responses in Acne Patients. J Invest Dermatol. 2014.
67. Sahdo B, Sarndahl E, Elgh F, Soderquist B. Propionibacterium acnes activates caspase-1 in human neutrophils. APMIS. 2013;121(7):652-63.
68. Li ZJ, Choi DK, Sohn KC, Seo MS, Lee HE, Lee Y, et al. Propionibacterium acnes Activates the NLRP3 Inflammasome in Human Sebocytes. J Invest Dermatol. 2014.
69. Agak GW, Qin M, Nobe J, Kim MH, Krutzik SR, Tristan GR, et al. Propionibacterium acnes Induces an IL-17 Response in Acne Vulgaris that Is Regulated by Vitamin A and Vitamin D. J Invest Dermatol. 2014;134(2):366-73.
70. Jasson F, Nagy I, Knol AC, Zuliani T, Khammari A, Dreno B. Different strains of Propionibacterium acnes modulate differently the cutaneous innate immunity. Exp Dermatol. 2013;22(9):587-92.
71. Lyte P, Sur R, Nigam A, Southall MD. Heat-killed Propionibacterium acnes is capable of inducing inflammatory responses in skin. Exp Dermatol.. 2009;18(12):1070-2.
72. Braga EG, Ananias RZ, Mussalem JS, Squaiella CC, Longhini AL, Mariano M, et al. Treatment with Propionibacterium acnes modulates the late phase reaction of immediate hypersensitivity in mice. Immunol Lett. 2003;88(2):163-9.
73. Tsuda K, Yamanaka K, Linan W, Miyahara Y, Akeda T, Nakanishi T, et al. Intratumoral injection of Propionibacterium acnes suppresses malignant melanoma by enhancing Th1 immune responses. PLoS One. 2011;6(12):e29020.
74. Reis VO, Silva JC, Souza GT, Semedo P, Buscariollo B, Pereira RL, et al. The polysaccharide fraction of Propionibacterium acnes modulates the development of experimental focal segmental glomerulosclerosis. Immunobiology. 2012;217(9):831-41.
75. Megid J, Peracoli MT, Curi PR, Zanetti CR, Cabrera WH, Vassao R, et al. Effect of bacillus of Calmette-Guerin, avridine and Propionibacterium acnes as immunomodulators on rabies in mice. Rev Inst Med Trop Sao Paulo. 1999;41(2):107-14.
76. Megid J, Peracolli MT, Curi PR, Zanetti CR, Cabrera WH, Vassao R, et al. Effect of vaccination and the immunomodulators "bacillus of Calmette-Guerin, avridine and Propionibacterium acnes" on rabies in mice. Comparative immunology, microbiology and infectious diseases. Comp Immunol Microbiol Infect Dis. 1998;21(4):305-18.
77. Iseki H, Takabatake N, Ota N, Ishigame T, Yokoyama N, Igarashi I. Babesia: the protective effects of killed Propionibacterium acnes on the infections of two rodent Babesia parasites in mice. Exp Parasitol. 2008;118(4):543-8.
78. Abel LC, Chen S, Ricca LG, Martins MF, Garcia M, Ananias RZ, et al. Adjuvant effect of LPS and killed Propionibacterium acnes on the development of experimental gastrointestinal nematode infestation in sheep. Parasite Immunol. 2009;31(10):604-12.
79. Silva JB, Oliveira SK, Campos IA, Carvalho-Junior CH, Coutinho Tda C, Silva TG. Propionibacterium acnes-killed attenuates the inflammatory response and protects mice from sepsis by modulating inflammatory factors. Braz J Infect Dis. 2013;17(1):20-6.
80. Kitagawa H, Yamanaka K, Kakeda M, Inada H, Imai Y, Gabazza EC, et al. Propionibacterium acnes vaccination induces regulatory T cells and Th1 immune responses and improves mouse atopic dermatitis. Experimental dermatology. 2011;20(2):157-8.
81. Paillot R. A systematic review of the immune-modulators Parapoxvirus ovis and Propionibacterium acnes for the prevention of respiratory disease and other infections in the horse. Vet Immunol Immunopathol. 2013;153(1-2):1-9.
82. Li L, Sun C, Yang F, Yang S, Feng X, Gu J, et al. Identification of proteins of Propionibacterium acnes for use as vaccine candidates to prevent infection by the pig pathogen Actinobacillus pleuropneumoniae. Vaccine. 2013;31(45):5269-75.
83. Fischer N, Mak TN, Shinohara DB, Sfanos KS, Meyer TF, Bruggemann H. Deciphering the intracellular fate of Propionibacterium acnes in macrophages. BioMed Res Int. 2013;2013:603046.
84. Debelian GJ, Olsen I, Tronstad L. Bacteremia in conjunction with endodontic therapy. Endod Dent Traumatol. 1995;11(3):142-9.
85. Park HJ, Na S, Park SY, Moon SM, Cho OH, Park KH, et al. Clinical significance of Propionibacterium acnes recovered from blood cultures: analysis of 524 episodes. J Clin Microbiol. 2011;49(4):1598-601.
86. Zhou Y, Hu Y, Li H. Role of Propionibacterium Acnes in Sarcoidosis: A Meta-analysis. Sarcoidosis, vasculitis, and diffuse lung diseases : official journal of WASOG / World Association of Sarcoidosis and Other Granulomatous Disorders. Sarcoidosis Vasc Diffuse Lung Dis.2013;30(4):262-7.
87. Yamada T, Eishi Y, Ikeda S, Ishige I, Suzuki T, Takemura T, et al. In situ localization of Propionibacterium acnes DNA in lymph nodes from sarcoidosis patients by signal amplification with catalysed reporter deposition. J Pathol. 2002;198(4):541-7.
88. Takemori N, Nakamura M, Kojima M, Eishi Y. Successful treatment in a case of Propionibacterium acnes-associated sarcoidosis with clarithromycin administration: a case report. J Med Case Rep. 2014;8(1):15.
89. Satoh F, Morita H, Tayama H, Inoue Y, Eishi Y, Yoshimura A. Renal sarcoidosis with limited lung manifestations expressing Propionibacterium acnes antigens in the affected tubulointerstitium. Am J Med Sci. 2013;346(3):250-2.
90. Negi M, Takemura T, Guzman J, Uchida K, Furukawa A, Suzuki Y, et al. Localization of propionibacterium acnes in granulomas supports a possible etiologic link between sarcoidosis and the bacterium. Mod Pathol. 2012;25(9):1284-97.
91. Homma JY, Abe C, Chosa H, Ueda K, Saegusa J, Nakayama M, et al. Bacteriological investigation on biopsy specimens from patients with sarcoidosis. Jpn J Exp Med. 1978;48(3):251-5.
92. Minegishi K, Aikawa C, Furukawa A, Watanabe T, Nakano T, Ogura Y, et al. Complete Genome Sequence of a Propionibacterium acnes Isolate from a Sarcoidosis Patient. Genome announcements. 2013;1(1).
93. Minami J, Eishi Y, Ishige Y, Kobayashi I, Ishige I, Kobayashi D, et al. Pulmonary granulomas caused experimentally in mice by a recombinant trigger-factor protein of Propionibacterium acnes. Journal of medical and dental sciences. 2003;50(4):265-74.
94. Kamata M, Tada Y, Mitsui A, Shibata S, Miyagaki T, Asano Y, et al. ICAM-1 deficiency exacerbates sarcoid-like granulomatosis induced by Propionibacterium acnes through impaired IL-10 production by regulatory T cells. Am J Pathol. 2013;183(6):1731-9.
95. Furusawa H, Suzuki Y, Miyazaki Y, Inase N, Eishi Y. Th1 and Th17 immune responses to viable Propionibacterium acnes in patients with sarcoidosis. Respiratory investigation. 2012;50(3):104-9.
96. De Marzo AM, Platz EA, Sutcliffe S, Xu J, Gronberg H, Drake CG, et al. Inflammation in prostate carcinogenesis. Nature Rev Cancer. 2007;7(4):256-69.
97. Cohen RJ, Shannon BA, McNeal JE, Shannon T, Garrett KL. Propionibacterium acnes associated with inflammation in radical prostatectomy specimens: a possible link to cancer evolution? J Urol. 2005;173(6):1969-74.
98. Fassi Fehri L, Mak TN, Laube B, Brinkmann V, Ogilvie LA, Mollenkopf H, et al. Prevalence of Propionibacterium acnes in diseased prostates and its inflammatory and transforming activity on prostate epithelial cells. Int J Med Microbiol. 2011;301(1):69-78.
99. Alexeyev OA, Marklund I, Shannon B, Golovleva I, Olsson J, Andersson C, et al. Direct visualization of Propionibacterium acnes in prostate tissue by multicolor fluorescent in situ hybridization assay. J Clin Microbiol. 2007;45(11):3721-8.
100. Bae Y, Ito T, Iida T, Uchida K, Sekine M, Nakajima Y, et al. Intracellular Propionibacterium acnes infection in glandular epithelium and stromal macrophages of the prostate with or without cancer. PLoS One. 2014;9(2):e90324.
101. Shannon BA, Cohen RJ, Garrett KL. Polymerase chain reaction-based identification of Propionibacterium acnes types isolated from the male urinary tract: evaluation of adolescents, normal adults and men with prostatic pathology. BJU international. 2006;98(2):388-92.
102. Shannon BA, Garrett KL, Cohen RJ. Links between Propionibacterium acnes and prostate cancer. Future oncology. 2006;2(2):225-32.
103. Shinohara DB, Vaghasia AM, Yu SH, Mak TN, Bruggemann H, Nelson WG, et al. A mouse model of chronic prostatic inflammation using a human prostate cancer-derived isolate of Propionibacterium acnes. Prostate. 2013;73(9):1007-15.
104. Mak TN, Yu SH, De Marzo AM, Bruggemann H, Sfanos KS. Multilocus sequence typing (MLST) analysis of Propionibacterium acnes isolates from radical prostatectomy specimens. Prostate. 2013;73(7):770-7.
105. Mak TN, Fischer N, Laube B, Brinkmann V, Metruccio MM, Sfanos KS, et al. Propionibacterium acnes host cell tropism contributes to vimentin- mediated invasion and induction of inflammation. Cell Microbiol. 2012;14(11):1720-33.
106. Severi G, Shannon BA, Hoang HN, Baglietto L, English DR, Hopper JL, et al. Plasma concentration of Propionibacterium acnes antibodies and prostate cancer risk: results from an Australian population-based case-control study. Br J Cancer. 2010;103(3):411-5.
107. Sperling JW, Kozak TK, Hanssen AD, Cofield RH. Infection after shoulder arthroplasty. Clin Orthop Relat Res. 2001(382):206-16.
108. Nodzo SR, Hohman DW, Crane JK, Duquin TR. Hemolysis as a Clinical Marker for Propionibacterium acnes Orthopedic Infection. Am J Orthop (Belle Mead NJ). 2014;43(5):E93-7.
109. Sperling JW, Cofield RH. Total shoulder arthroplasty after attempted shoulder arthrodesis: report of three cases. Journal of shoulder and elbow surgery. J Shoulder Elbow Surg. 2003;12(3):302-5.
110. Bemer P, Corvec S, Tariel S, Asseray N, Boutoille D, Langlois C, et al. Significance of Propionibacterium acnes-positive samples in spinal instrumentation. Spine (Phila Pa 1976). 2008;33(26):E971-6.
111. Bayston R, Ashraf W, Barker-Davies R, Tucker E, Clement R, Clayton J, et al. Biofilm formation by Propionibacterium acnes on biomaterials in vitro and in vivo: impact on diagnosis and treatment. J Biomed Mater Res A. 2007;81(3):705-9.
112. Mhaidli HH, Der-Boghossian AH, Haidar RK. Propionibacterium acnes delayed infection following spinal surgery with instrumentation. Musculoskeletal Surg. 2013;97(1):85-7.
113. Kurz M, Kaufmann BA, Baddour LM, Widmer AF. Propionibacterium acnes prosthetic valve endocarditis with abscess formation: a case report. BMC Infect Dis. 2014;14:105.
114. Beliaev AM, Roberts SA, Pemberton J, Haydock DA. Propionibacterium acnes biofilm endocarditis requiring radical cardiac debridement and prosthetic valve replacements. ANZ J Surg. 2014 Feb 17. [Epub ahead of print].
115. Guio L, Sarria C, de Las Cuevas C, Gamallo C, Duarte J. Chronic Prosthetic Valve Endocarditis Due to Propionibacterium acnes: An Unexpected Cause of Prosthetic Valve Dysfunction. Rev Esp Cardiol (Engl Ed). 2009;62(2):167-77.
116. Mirdha BR, Kumar P. Primary anaerobic bacterial meningitis caused by Propionibacterium acnes. Postgrad Med J. 1993;69(812):499-500.
117. Ueunten D, Tobias J, Sochat M, Miranda C, Mulligan M, Yoshikawa TT. An unusual cause of bacterial meningitis in the elderly. Propionibacterium acnes. Archives Neurol. 1983;40(6):388-9.
118. Schlesinger JJ, Ross AL. Propionibacterium acnes meningitis in a previously normal adult. Arch Intern Med. 1977;137(7):921-3.
119. French RS, Ziter FA, Spruance SL, Smith CB. Chronic meningitis caused by Propionibacterium acnes. A potentially important clinical entity. Neurology. 1974;24(7):624-8.
120. Zaffiri L, Abdulmassih R, Boyaji S, Bagh I, Campbell AR, Loehrke ME. Brain abscess induced by Propionibacterium acnes in a patient with severe chronic sinusitis. New Microbiol. 2013;36(3):325-9.
121. Adesina OO, Stagg BC, Digre KB, Katz BJ, Quigley EP, Palmer CA, et al. Optic Neuropathy Caused by Propionibacterium acnes Pachymeningitis. J Neuroophthalmol. 2014;34(3):264-7.
122. Aubin GG, Portillo ME, Trampuz A, Corvec S. Propionibacterium acnes, an emerging pathogen: From acne to implant-infections, from phylotype to resistance. Med Mal Infect. 2014;44(6):241-50.
123. Shailaja S, Kamath Y, Hazarika M, Vishwanath S. Acute post-traumatic endophthalmitis secondary to Propionibacterium acnes. BMJ Case Rep. 2013;2013.
124. Uckay I, Dinh A, Vauthey L, Asseray N, Passuti N, Rottman M, et al. Spondylodiscitis due to Propionibacterium acnes: report of twenty-nine cases and a review of the literature. Clin Microbiol Infect. 2010;16(4):353-8.
125. Harris AE, Hennicke C, Byers K, Welch WC. Postoperative discitis due to Propionibacterium acnes: a case report and review of the literature. Surgical Neurol. 2005;63(6):538-41; discussion 41.
126. Hernandez-Palazon J, Puertas-Garcia JP, Martinez-Lage JF, Tortosa JA. Lumbar spondylodiscitis caused by Propionibacterium acnes after epidural obstetric analgesia. Anesth Analg. 2003;96(5):1486-8, table of contents.
127. Retornaz F, Roche PH, Seux V, Caperan C, Touta A, Soubeyrand J. [Propionibacterium acnes spondylodiscitis: a case report]. Rev Med Interne. 2001;22(2):199-200.
128. Hammann C, Dudler J, Gaumann U, Landry M, Gerster JC. [Spondylodiscitis due to Propionibacterium acnes. Case report and review of the literature]. Schweiz Med Wochenschr. 1999;129(40):1456-60.
129. Chia JK, Nakata MN. Intervertebral diskitis caused by Propionibacterium acnes: a report of four cases. Clin Infect Dis. 1996;23(3):643-4.
130. Burki F, Treves R, Desproges-Gotteron R, Denis F. [A case of spondylodiscitis caused by Propionibacterium acnes and Peptococcus constellatus]. Revue du rhumatisme et des maladies osteo-articulaires. 1983;50(7):541-3.
131. Mesado D, Sarria C, Bustamante J, Rodriguez JE, Dominguez L, Olivera MJ. Constrictive Infectious Pericarditis Caused by Propionibacterium acnes. Rev Esp Cardiol (Engl Ed). 2013;66(5):407-9.
132. Harlock JA, Qadura M, Lee G, Szalay DA. Infected aortic stent graft with Propionibacterium acnes. Vasc Endovascular Surg. 2013;47(5):394-6.
133. Pozo E, Vilacosta I, Canadas MV, Del Trigo M, Silva J, Rodriguez E. Chronic infective endarteritis due to Propionibacterium acnes on aortic prosthetic graft. Rev Esp Cardiol (Engl Ed). 2012;65(2):194-5.
134. Hayashi Y, Eguchi H, Miyamoto T, Inoue M, Mitamura Y. A Case of Delayed-Onset Propionibacterium acnes Endophthalmitis after Cataract Surgery with Implantation of a Preloaded Intraocular Lens. Case reports in ophthalmology. 2012;3(3):291-7.
135. Levitt MR, Gabikian P, Pottinger PS, Silbergeld DL. Propionibacterium acnes osteomyelitis occurring 23 years after craniotomy: case report and review of literature. Neurosurgery. 2011;69(3):E773-9; discussion E9.
136. Asseray N, Papin C, Touchais S, Bemer P, Lambert C, Boutoille D, et al. Improving diagnostic criteria for Propionibacterium acnes osteomyelitis: a retrospective analysis. Scand J Infect Dis. 2010;42(6-7):421-5.
137. Kormondi S, Terhes G, Pinter S, Urban E. Granulomatous Propionibacterium acnes infection after trauma surgery. Anaerobe. 2011;17(5):259-61.
138. Niazi SA, Clarke D, Do T, Gilbert SC, Mannocci F, Beighton D. Propionibacterium acnes and Staphylococcus epidermidis isolated from refractory endodontic lesions are opportunistic pathogens. J Clin Microbiol. 2010;48(11):3859-69.
139. Ovodenko B, Seedor JA, Ritterband DC, Shah M, Yang R, Koplin RS. The prevalence and pathogenicity of Propionibacterium acnes keratitis. Cornea. 2009;28(1):36-9.
140. Pandya AG. Top accessed article: propionibacterium acnes and the pathogenesis of progressive macular hypomelanosis. Arch Dermatol. 2012;148(11):1256.
141. de Morais Cavalcanti SM, de Franca ER, Magalhaes M, Lins AK, Brandao LC, Magalhaes V. A quantitative analysis of Propionibacterium acnes in lesional and non-lesional skin of patients with progressive macular hypomelanosis by real-time polymerase chain reaction. Braz J Microbiol. 2011;42(2):423-429.
142. Cavalcanti SM, de Franca ER, Lins AK, Magalhaes M, de Alencar ER, Magalhaes V. Investigation of Propionibacterium acnes in progressive macular hypomelanosis using real-time PCR and culture. Int J Dermatol. 2011;50(11):1347-52.
143. Wang E, Lee JS, Hee TH. Is propionibacterium acnes associated with hair casts and alopecia? International journal of trichology. 2012;4(2):93-7.
144. Colina M, Lo Monaco A, Khodeir M, Trotta F. Propionibacterium acnes and SAPHO syndrome: a case report and literature review. Clin Exp Rheumatol. 2007;25(3):457-60.
145. Hall GS, Pratt-Rippin K, Meisler DM, Washington JA, Roussel TJ, Miller D. Growth curve for Propionibacterium acnes. Curr Eye Res. 1994;13(6):465-6.
146. Perry A, Lambert P. Propionibacterium acnes: infection beyond the skin. Expert Rev Anti Infect Ther. 2011;9(12):1149-56.
147. Butler-Wu SM, Burns EM, Pottinger PS, Magaret AS, Rakeman JL, Matsen FA, 3rd, et al. Optimization of periprosthetic culture for diagnosis of Propionibacterium acnes prosthetic joint infection. J Clin Microbiol. 2011;49(7):2490-5.
148. Poppert S, Riecker M, Essig A. Rapid identification of Propionibacterium acnes from blood cultures by fluorescence in situ hybridization. Diagn Microbiol Infect Dis. 2010;66(2):214-6.
149. Alexeyev OA, Jahns AC. Sampling and detection of skin Propionibacterium acnes: current status. Anaerobe. 2012;18(5):479-83.
150. Martin WJ, Gardner M, Washington JA, 2nd. In vitro antimicrobial susceptibility of anaerobic bacteria isolated from clinical specimens. Antimicrob Agents Chemother. 1972;1(2):148-58.
151. Ross JI, Eady EA, Carnegie E, Cove JH. Detection of transposon Tn-5432-mediated macrolide-lincosamide-streptogramin B (MLSB) resistance in cutaneous propionibacteria from six European cities. J Antimicrob Chemother. 2002;49(1):165-8.
152. El-Mahdy TS, Abdalla S, El-Domany R, Mohamed MS, Ross JI, Snelling AM. Detection of a new erm(X)-mediated antibiotic resistance in Egyptian cutaneous propionibacteria. Anaerobe. 2010;16(4):376-9.
153. Furustrand Tafin U, Trampuz A, Corvec S. In vitro emergence of rifampicin resistance in Propionibacterium acnes and molecular characterization of mutations in the rpoB gene. J Antimicrob Chemother. 2013;68(3):523-8.
154. Lomholt HB, Kilian M. Clonality and Anatomic Distribution on the Skin of Antibiotic Resistant and Sensitive Propionibacterium acnes. Acta Derm Venereol. 2014;94(5):534-8.
155. Ross JI, Snelling AM, Carnegie E, Coates P, Cunliffe WJ, Bettoli V, et al. Antibiotic-resistant acne: lessons from Europe. Br J Dermatol. 2003;148(3):467-78.
156. Coates P, Vyakrnam S, Eady EA, Jones CE, Cove JH, Cunliffe WJ. Prevalence of antibiotic-resistant propionibacteria on the skin of acne patients: 10-year surveillance data and snapshot distribution study. Br J Dermatol. 2002;146(5):840-8.
157. Oprica C, Emtestam L, Lapins J, Borglund E, Nyberg F, Stenlund K, et al. Antibiotic-resistant Propionibacterium acnes on the skin of patients with moderate to severe acne in Stockholm. Anaerobe. 2004;10(3):155-64.
158. Kurokawa I, Nishijima S, Kawabata S. Antimicrobial susceptibility of Propionibacterium acnes isolated from acne vulgaris. Eur J Dermatol. 1999;9(1):25-8.
159. Nakase K, Nakaminami H, Noguchi N, Nishijima S, Sasatsu M. First report of high levels of clindamycin-resistant Propionibacterium acnes carrying erm(X) in Japanese patients with acne vulgaris. J Dermatol. 2012;39(9):794-6.
160. Schafer F, Fich F, Lam M, Garate C, Wozniak A, Garcia P. Antimicrobial susceptibility and genetic characteristics of Propionibacterium acnes isolated from patients with acne. Int J Dermatol. 2013;52(4):418-25.
161. Mendoza N, Hernandez PO, Tyring SK, Haitz KA, Motta A. Antimicrobial susceptibility of Propionibacterium acnes isolates from acne patients in Colombia. Int J Dermatol. 2013;52(6):688-92.
162. Gonzalez R, Welsh O, Ocampo J, Hinojosa-Robles RM, Vera-Cabrera L, Delaney ML, et al. In vitro antimicrobial susceptibility of Propionibacterium acnes isolated from acne patients in northern Mexico. Int J Dermatol. 2010;49(9):1003-7.
163. Luk NM, Hui M, Lee HC, Fu LH, Liu ZH, Lam LY, et al. Antibiotic-resistant Propionibacterium acnes among acne patients in a regional skin centre in Hong Kong. J Eur Acad Dermatol Venereol. 2013;27(1):31-6.
164. Abdel Fattah NS, Darwish YW. In vitro antibiotic susceptibility patterns of Propionibacterium acnes isolated from acne patients: an Egyptian university hospital-based study. J Eur Acad Dermatol Venereol. 2013;27(12):1546-51.
165. Toyne H, Webber C, Collignon P, Dwan K, Kljakovic M. Propionibacterium acnes (P. acnes) resistance and antibiotic use in patients attending Australian general practice. Australas J Dermatol. 2012;53(2):106-11.
166. Oprica C, Nord CE, Bacteria ESGoARiA. European surveillance study on the antibiotic susceptibility of Propionibacterium acnes. Clin Microbiol Infect. 2005;11(3):204-13.
167. Wang B, Toye B, Desjardins M, Lapner P, Lee C. A 7-year retrospective review from 2005 to 2011 of Propionibacterium acnes shoulder infections in Ottawa, Ontario, Canada. Diagn Microbiol Infect Dis. 2013;75(2):195-9.
168. Furustrand Tafin U, Corvec S, Betrisey B, Zimmerli W, Trampuz A. Role of rifampin against Propionibacterium acnes biofilm in vitro and in an experimental foreign-body infection model. Antimicrob Agents Chemother. 2012;56(4):1885-91.
169. Crane JK, Hohman DW, Nodzo SR, Duquin TR. Antimicrobial susceptibility of Propionibacterium acnes isolates from shoulder surgery. Antimicrob Agents Chemother. 2013;57(7):3424-6.
This study was carried out at the Dermatology Department, Faculdade de Ciências Médicas de Minas Gerais - Belo Horizonte (MG), Brazil.
 All content the journal, except where identified, under the Creative Commons Attribution 4.0 International licence - ISSN-e 1984-8773
All content the journal, except where identified, under the Creative Commons Attribution 4.0 International licence - ISSN-e 1984-8773



 Read in Portuguese
Read in Portuguese
 Portuguese PDF
Portuguese PDF
 Print
Print
 Send this article by email
Send this article by email
 How to cite this article
How to cite this article
 Submit a comment
Submit a comment
 Mendeley
Mendeley
 Pocket
Pocket
{kind=link}
{kind=link}